Interdisciplinary Ocean Science is Evolving
by James J. McCarthy1, Allan R. Robinson1 and Brian J. Rothschild2
The earthÆs surface fluids comprise a dynamic and highly variable coupled physical, biological and chemical system. The oceanic component of this system is controlled by ecosystem-biogeochemical-physical processes interacting over a vast range of scales in space and time. This is a propitious time in ocean science. The large-scale global studies of physical, biogeochemical, and biological ocean phenomena and processes now underway are setting the stage for a new generation of interdisciplinary ocean studies. Moreover, an opportunity exists to set future ocean studies efficiently on wide-ranging temporal and spatial scales in the context of satellite and in situ sensing systems that are now being planned and developed. Two decades of progress in describing, modeling and forecasting of the physical synoptic-mesoscale features in the ocean now makes it feasible to provide mesoscale fields over large-scale domains suitable for realistic physical-biological interactive process studies. Thus the substantial fundamental interdisciplinary problems of ocean science, long recognized as of great importance, are now tractable. As the scope of interdisciplinary ocean science expands, its complexity enlarges and its research intensifies, ocean science is undergoing a conceptual evolution.
An overarching fundamental interdisciplinary question has focused much effort in biological oceanography: How are growth and reproduction of organisms in the sea, i.e. biological production, regulated, allocated amongst different groups of organisms, and processed to allow either for sustained local production through recycling or for export? Aspects of this question have been pursued with great success in the last few decades using both disciplinary and some interdisciplinary approaches. Such aspects include: the quantitative relationship between radiant energy and rate of photosynthesis; the roles of macro and micro nutrients in determining both the rate of primary production and the qualitative character of the phytoplankton assemblage; the relative importance of different taxa and size classes of phytoplankton and zooplankton in determining food web relationships and the partitioning between new and recycled production; and the expanded role of microbes both as consumers of dissolved organic carbon and as prey for micro-zooplankton (e.g. Falkowski and Woodhead, 1992). New sophistication of understanding regarding biological processes must, however, be set in the appropriate physical context in order to be fully applicable to the general question stated above. During the last two decades research addressing jointly and realistically the physical, biological and biogeochemical aspects of ocean circulation features has revealed new quantitative understanding regarding the regulation of nutrient supply, the processing and retention of nutrient material by organisms, and the distribution of organisms (e.g. Joyce and Wiebe, 1992). Systematic extension and evolution of these and other interdisciplinary successes now offers real promise for elucidating societally important aspects of the overarching question, such as variability in stocks of marine species harvested for human use and the oceanÆs capacity to store carbon under different climate regimes.
Our thesis is that in order to be successful large-scale studies of marine ecosystems must be carried out in a context that balances the perspectives of physics, population dynamics, and biogeochemical processes. An increasing number of ocean scientists recognize that the traditional subdisciplines of ocean science no longer constitute a sufficient framework for advancing ocean scientific knowledge. It is necessary to study the dynamic mechanisms by which physical processes influence biogeochemical and population-dynamic processes, their interactions and attendant feedbacks. These studies must attain a new level of understanding as to how mesoscale and smaller scale processes and interactions determine the distributions and variabilities of biological and chemical variables over a wide variety of temporal and spatial scales, including larger scales. Ecosystems are comprised of a seemingly infinite number of state variables for the diverse species, life-stages, nutrients, etc. Thus, it must be recognized that alternative small dimensionality representations may lead to different qualitative and quantitative conclusions regarding how physical forcing affects the population-dynamic and biogeochemical processes and hence the temporal-spatial variability of the biology and chemistry. Robust small dimensionality representations must be sought and achieved.
The tractability of these interdisciplinary problems requires a systems approach for efficient and cost-effective research. Elements of the research system include: a versatile set of linked physical-biological-chemical models coupled to a flexible multiplatform, multisensor observational network via interdisciplinary data assimilation schemes; a sampling and sensor optimization module for observational network design via theory and simulations; and general analysis schemes for the identification and interpretation of physical, biogeochemical and ecosystem dynamical processes. Because of the ranges of requisite scales, matched nests of models and sampling schemes are necessary.
Challenges and Issues
In the late 1950s to early 1960s, biological oceanography became process oriented with the development of dynamic models of variability and productivity of the plankton, and with new techniques such as: a suite of isotopic techniques for quantifying plankton production rates; new microscopic, chemical and electronic means for assessing both the abundances and the roles of different size classes and taxa of phyto-, zoo-, and bacterio-plankton in the overall flow of energy and material throughout the plankton community. In the 1980s the advent of remote sensing, new computing capabilities, and disciplinary maturity allowed a surge in interdisciplinary field and modeling approaches to subbasin scales of physical-biological dynamical studies of coastal and open ocean waters.
As interdisciplinary research in ocean science began to flourish, failures of traditional disciplinary approaches with respect to stock size predictions for marine species of commercial interest were becoming increasingly common. While the role of physics in fish population biology has long been recognized, the fully interactive physical-ecological-biogeochemical nature of central and critical relationships was not fully realized, and methods to study it with adequate detail did not yet exist. In addition, the entire earth science community has, in this same time frame, become aware of a profound change with uncertain implications for future climate: the secular increase in annually averaged atmospheric carbon dioxide content. Predictions of future atmospheric carbon dioxide content remain in doubt. In part, this results from uncertainty in forecasts relating to the strength of the anthropogenic source term. But there are also serious questions regarding the oceanÆs response to this perturbation in the global carbon cycle. Coupled atmosphere-ocean general circulation models, when forced with altered radiative physics consistent with projections for future emissions of CO2 and releases of other so-called greenhouse gases, yield scenarios with different, in some cases dramatically different, ocean circulation (Manabe and Stouffer, 1993). Changes in the physical components of the climate system will influence biological and biogeochemical ocean processes, and together this ensemble of climate system processes determines the oceanÆs capacity to store carbon (Siegenthaler and Sarmiento, 1993).
Developments in Ocean Physics: Physical processes and phenomena which may affect biological and chemical processes significantly include: turbulence, fine and microstructure, internal waves, tides and mixing; surface and bottom boundary layer daily, seasonal, storm induced and (sub)mesoscale forced transports and variabilities; synoptic/mesoscale fronts associated with meandering currents, cast off ring vortices and mid-ocean eddies, jet filaments and submesoscale lenses; coastal upwelling, shelf-break and inshore fronts, coastal along-shore and cross-shore currents, transports and mixing; general circulation, seasonal cycles, inter- and multi-annual anomalies and climate drift. The most energetic variability phenomena in the oceans are synoptic/mesoscale (10° -102 days; 10° -102 km) (Robinson, 1983) which encompass the life-times and life-time spatial ranges of most organisms in the sea. While globally, it is the large scale and average effects of the mesoscale which are of primary interest biologically, explicit mesoscale resolution is still required in order to research mesoscale interdisciplinary interactions.
Physical oceanography has made considerable progress in the last two decades in both descriptive and dynamical areas. New measurement techniques, instruments and platforms together with adequate space-time sampling strategies have provided a new data base that reveals the typical synoptic circulation structures of the energetic mesoscale variability. These include the mesoscale eddies, planetary and equatorial waves, fronts, current meanders and filaments which can significantly affect biogeochemical cycles and ecosystems dynamics. The evolution of these mesoscale structures is punctuated by intermittent and rapid energetic events, such as the necking-off of a ring eddy from a current or the merging of two eddies. Nonlinear internal dynamical processes, analogous to processes which cause weather and storms in the atmosphere, govern the oceanic mesoscale. Mesoscale dynamical studies and model developments have resulted in new numerical dynamical ocean models which adequately represent the basic physics of the mesoscale phenomena and which are suitable for use with real ocean data over large scale regions. Additionally, substantial progress is occurring in describing the structure and dynamics of the global general circulation, regional circulations and the formation of deep and intermediate water masses. Thus, knowledge now exists to provide field estimates required for interdisciplinary modeling and research. Such research needs to identify the horizontal and vertical transport mechanisms for biological material specific to various regions and processes and to postulate general paradigms. Feedback mechanisms from the biology to the physics also require investigation. Examples of the latter are the biooptical effects in the upper ocean and bioconvection, i.e., Rayleigh-Benard convection which arises from biologically induced density differences.
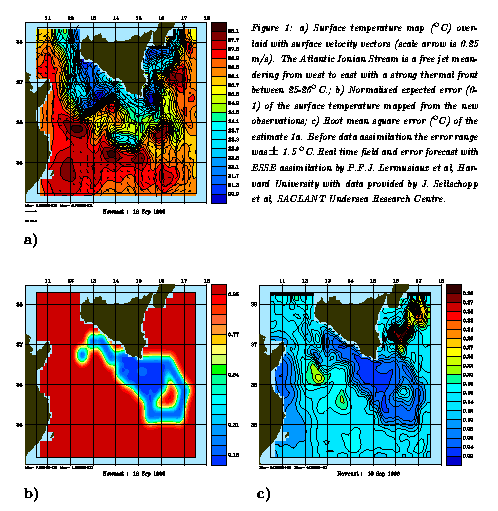
Data Assimilation and Field Estimation: Dynamical models are a powerful vehicle for conveying physical oceanographic information for scientific and practical applications. Space-time distributions of state variables in the ocean are called fields. The resource requirements for obtaining accurate large regional scale three- and four-dimensional maps and time series of fields with mesoscale resolution via direct sampling on an observational grid are generally prohibitive. Data assimilation, which melds observations with dynamics, provides the only feasible basis for obtaining accurate synoptic mesoscale realizations over the space-time scales and domains of interest. Data assimilation dynamically adjusts and dynamically interpolates data inserted into models. There is a duration of predictability for atmospheric and oceanic forecasts which is related to the nonlinear transfer of errors and noise into scales of interest. Beyond the predictability limit the forecast will still contain realistic events and structures but not those actually occurring at that time in nature. Loss of predictability can be controlled by sequential updating of the model forecast with new observations. The relative weights of the data and the forecast when the observations are melded with dynamics are based on estimates of both observational errors and model errors. Thus error models are an intrinsic element of data assimilation schemes, and errors are propagated together with the forecast fields. This process is illustrated in Figure 1 with a real time physical forecast for the Straits of Sicily and the Ionian Sea. The domain is 400 km by 600 km in extent, eddies and meanders occur on scales from 10 to 100 km, and grids were nested with resolutions of 1, 3, 9 and 27 kms. The region was initialized in real time on shipboard with hydrographic data taken from the ship and subsequently updated by temperature vs. depth probes dropped by aircraft. Figure 1a shows the melded estimate after assimilation, Figure 1b shows the expected error of the new data assimilated, and Figure 1c maps the error associated with the estimated field of Figure 1a. Other applications of data assimilation or inverse methods allow for the estimation of parameters such as eddy diffusivities and rate parameters and also for the inference of processes from the balances of terms in dynamical equations (Malanotte-Rizzoli, 1996). A most important aspect of current research is the extension of physical oceanographic data assimilation and simulation methods to biological and chemical data sets and ocean biogeochemical/ ecosystem models. Such extensions are now underway and should soon produce sophisticated interdisciplinary field estimates such as illustrated for the physics in Figure 1.
Figure 1 - a real time physical forecast for the Straits of Sicily and the Ionian Sea: a) the melded estimate after assimilation; b) the expected error of the new data assimilated, and c) the error associated with the estimated field of Figure 1a.
Plankton Biogeochemical Cycles and Physics: The distribution of organisms in the sea is strongly influenced by the horizontal pattern of ocean circulation. This is true for the plankton and also true for many species of nekton whose faunal boundaries are defined by circulation patterns. Upwelling to the upper ocean and mixing, when sufficiently intense to enrich near-surface waters with nutrients, influence the distribution, abundance, and growth rates of plankton.
Plankton "blooms" result from enhanced growth and/or retention of phytoplankton in the euphotic zone. Studies over the last several decades have provided a rich source of data relating to the temporal and spatial extent of the spring bloom for plankton in many coastal regions. And yet, factors responsible for timing, spatial extent, and duration of such bloom events are still poorly documented for many coastal regions. In deep water, results of sediment trap studies show that during bloom periods the flux of particulate organic matter to deep water and the ocean floor is enhanced. Results of one of the most complex models to date that marries a multiple compartment nitrogen-based ecosystem model with a non-eddy resolving three-dimensional seasonal general circulation model (Sarmiento et al., 1993) demonstrate sensitivity to both nitrate availability and zooplankton dynamics in regulating rates of primary production and levels of biomass. Results from a similar model, applied to a limited region of the northeast Atlantic, but with fewer compartments and the ability to resolve mesoscale eddies, rationalizes the 1989 spring bloom and indicates an important post-bloom nutrient injection mechanism into the upper ocean by mesoscale eddy interactions (McGillicuddy et al., 1995).
There is at this time an increasing awareness of the need to document with greater spatial and temporal detail the distributions of marine plankton in general, and key groups in particular, in order to understand better the role of these organisms in the coupling of global biogeochemical cycles and the physical climate system (Evans and Fasham, 1993). Moreover, it is now widely accepted that in order to quantify the influence of physical processes on ocean biology, biological sampling must be matched to the appropriate temporal and spatial scales for relevant physical properties and processes.
Food Web Linkages: Food webs represent the flow of biological energy among the diverse populations in an ecosystem. In the past two decades the concept of the food web, with attendant bifurcations, loops and multidirectional flows of material, has replaced the simpler earlier paradigm of a linear food chain. At the base of a food web, phytoplankton transform the sunÆs energy, dissolved nutrients, and CO2 into phytoplankton biomass. The phytoplankton are grazed upon by herbivorous zooplankton. The phytoplankton also exude organic molecules, which are consumed by bacteria. The bacteria are eaten by microzooplankton. The microzooplankton and the herbivorous zooplankton are fed upon by larger zooplankton and so on. Physical forcing modulates the efficiency of energy transfers from one component of the food web to another component of the food web. To name a few examples, these processes control: the rate of primary production by influencing the exposure of phytoplankton to light and remineralized nutrients; the rates of growth and mortality of all organisms because of the important effect of temperature on metabolism; and the rates of encounter between predator and prey. There is little doubt that variability in the food web structure itself (i.e., its size and complexity) is modified by, and interacts with, physical forcing (Rothschild, 1988) to affect the efficiency of primary production and the transfer of biological energy from one population in the food web to the next, eventually terminating in the metabolism of the largest predators. Laboratory and field studies have clarified these pathways and added valuable information regarding population level specifications in rates for these transformations in many marine ecosystems (Harris, 1995). However, critical gaps in understanding remain.
An understanding of observed population variability depends upon an understanding of the factors that influence the composition of the plankton community. Planktonic ecosystems vary with regard to turnover rates. At one end of the spectrum are communities dominated by small phytoplankton that are grazed primarily by protozoans with extensive recycling via the microbial loop, with a large fraction of the carbon photosynthetically fixed rapidly respired back to CO2 with turnover times of hours to days. At the other end are communities in which large phytoplankton cells, such as diatoms, are subjected to low grazing pressure, and gravitationally settle to the deep ocean. Under these conditions a significant portion of the carbon photosynthetically fixed by phytoplankton is held in the deep ocean on time scales of the ocean circulation, seasons to millennia. Most natural ecosystems lie between these two extremes.
Several factors influence the composition of the plankton community. Physical mixing and transport processes contribute both directly and indirectly. Large diatoms require higher rates of upper ocean mixing to remain in suspension than do small diatoms, while motile flagellated phytoplankton require none. A pulse of nutrients to the upper ocean can stimulate a plankton bloom, and the nitrogen to silicon nutrient ratio, and iron availability, will influence the composition of the bloom community.
Advancing our understanding of food-web structure requires addressing the problem at appropriate levels of complexity. The structure of the food web is relatively well known in the context of a more or less simple linear structure, for which various sizes of plankton exhibit fairly well understood trophic functioning. However, rates of material and energy flux among the components of major food webs are poorly known. Most are conjectured or derived from limited experiments so, at best, the inferences that can be drawn from their magnitude, especially in a highly variable ocean, are limited. Even more problematic are the effects of physical forcing on food web structure and functions. Advancing our understanding of pathways, the regulation of rates, and feedbacks implied by food-web structure, population dynamics, and the role of physical forcing in driving variability in the biodynamics of the ocean will require advanced and novel interdisciplinary models to serve as constraints in data assimilation schemes.
Applications of the New Science
Ocean science and technology have long served societyÆs needs. Applications in weather prediction, utilization of mineral and biotic resources, maritime and naval operations and coastal management, are but a few common examples. However, many such successes are typically based primarily within a single scientific discipline. While some ocean problems of societal interest will continue to benefit from this approach, others require a new, truly interdisciplinary approach. Examples of such problems introduced above, are variability in stocks of marine species harvested for human use and the oceanÆs capacity to store carbon under different climate regimes.
In 1972 the Peruvian anchovetta fishery, which at that time had the highest annual yield of any marine fishery, collapsed with severe consequences for the economy of Peru. After two decades the population is increasing. However, debate continues regarding the respective roles of the 1972 El Nino, excessive harvest, and other factors in causing this major adjustment of a marine ecosystem. Subsequent analysis of the biological data associated with the Humboldt Current indicates that the collapse of the anchovetta fishery was paralleled with a reduction in zooplankton abundance. Interestingly, the abundance of large zooplankton in the California Current has continued to decline in the four decades since the collapse of the sardine fishery in these waters. It is increasingly apparent that stock size for both anchovy and sardine populations inhabiting the Pacific Ocean boundary currents vary on a wide range of time scales throughout this ocean basin, and the paleo record indicates that this is not a recent phenomenon. While fishing pressure may have accelerated the declines of some of these stocks, in the case of the Humboldt Current, the sustained low abundances of both the anchovetta and the zooplankton cannot be explained solely by fishing pressure or recent El Nino events.
Fluxes of carbon among the atmosphere, terrestrial and marine components of the EarthÆs carbon cycle have strong implications for climate. A change in climate and upper ocean physics could by various scenarios induce feedback to atmospheric CO2 content via biological processes. These terrestrial and marine components of the physical and biogeochemical systems that regulate the CO2 content of the atmosphere do not presently accommodate more than half of the current anthropogenic source from fossil fuel combustion and changes in land use and land cover. What determines the capacity of terrestrial and ocean systems to sequester this CO2 is presently unknown, and for the ocean surely will not be resolved without explicit consideration of the large scale consequences of mesoscale processes (Heimann, 1993).
Both of these examples reinforce the point that the Earth system consists of dynamic and highly variable physical, chemical, and biological components. It is clear that contemporary interactions within this system influence both the cycles of dominance for marine organisms and the nature of the climate system broadly defined. The ocean-atmosphere aspects are of particular scientific interest because as the record of the past demonstrates, these interacting ecosystem-biogeochemical-physical processes were responsible for modifying the composition of the EarthÆs atmosphere and ocean to allow life as we know it to evolve (Kasting, 1993).
Implementing the New Science: The Systems Approach
Research which stimulates an understanding of ocean processes that span the traditional disciplinary domain of the sciences, and which focuses upon the synthesis among the large data sets now being gathered pertaining to biological and physical variability in the ocean, is intimately linked to the conceptual basis for the next generation of global ocean observing systems. The evolution of interdisciplinary ocean science involves an hierarchy of complex coupled physical-biogeochemical-ecosystem dynamical processes, with nonlinearities and feedbacks which span a multitude of scales in time and space. The formidable and challenging research tasks required to address the related hierarchy of scientific problems and to move the science forward as rapidly as necessary is feasible, but only by the systems approach. The systems concept involves novel and advanced models assimilating on nested scales matched biological and physical data sets, acquired from multiplatform and multisensor observational networks. Quantitative sampling theoretic methods embedded within observational system simulation experiments must be utilized for network design and to set a priori efficiency and accuracy requirements. Such systems are the only way to achieve the simulation, predictive and monitoring capabilities needed for the research, development and management of the oceanic component of the whole earth system.
Acknowledgments
We are grateful to several colleagues for helpful comments and discussions. Research that stimulated the dialogue that resulted in this essay was supported by NASA, NOAA, NSF and ONR.
References
Evans, G.T. and M.J.R. Fasham, eds., 1993: Towards a Model of Ocean Biogeochemical Processes, NATO ASI Series, Series I: Global Environmental Change, Vol. 10, 350 pp., Springer-Verlag, Germany.
Falkowski, P.G. and A.D. Woodhead, eds., 1992: Primary Productivity and Biogeochemical Cycles in the Sea, Environmental Science Research, 43, 550 pp., Plenum Press, New York.
Harris, R., ed., 1995: Zooplankton Production, ICES Journal of Marine Science, 52, Nos. 3 and 4, 773 pp., Academic Press, London.
Heimann, M., ed., 1993: The Global Carbon Cycle, NATO ASI Series, Series I: Global Environmental Change, Vol. 15, 599 pp., Springer-Verlag, Germany.
Joyce, T.M. and P.H. Wiebe, eds., 1992: Warm-Core Rings: Interdisciplinary Studies of Kuroshio and Gulf Stream Rings, Deep Sea Research, 39, Supplementary Issue No. 1A March 1992, 417 pp., Pergamon Press, New York.
Kasting, J.F., 1993: EarthÆs Early Atmosphere, Science, 259: 920-926.
Malanotte-Rizzoli, P., ed., 1996: Modern Approaches to Data Assimilation in Ocean Modeling, Elsevier Science, The Netherlands, 455 pp.
Manabe, S. and R.J. Stouffer, 1993: Century-scale Effects of Increased Atmospheric Carbon Dioxide on the Ocean-Atmosphere System, Nature, 364: 215-218.
McGillicuddy, Jr., D.J., A.R. Robinson and J.J. McCarthy, 1995: Coupled physical and biological modeling of the spring bloom in the North Atlantic (II): Three-dimensional bloom and post-bloom effects, Deep-Sea Research I, 42(8), 1359-1398.
Robinson, A.R., ed., 1983: Eddies in Marine Science, Springer-Verlag, Germany, 609 pp.
Rothschild, B.J., ed., 1988: Toward a Theory on Biological-Physical Interactions in the World Ocean, Series C: Mathematical and Physical Sciences, Vol. 239, 650 pp., Kluwer Academic Publishers, The Netherlands.
Sarmiento, J.L., R.D. Slater, M.J.R. Fasham, H.W. Ducklow, J.R. Toggweiler and G.T. Evans, 1993: A seasonal three-dimensional ecosystem model of nitrogen cycling in the North Atlantic euphotic zone, Global Biogeochem. Cycles, 7(2): 415-450.
Siegenthaler, U. and J.L. Sarmiento, 1993: Atmospheric Carbon Dioxide and the Ocean, Nature, 365: 119-125.